During development multiple transcriptional and post-transcriptional networks are coordinately regulated to drive organ maturation, tissue formation, and cell fate. Particularly interesting is the fact that more than 90% of the human genes undergo alternative splicing. Alternative splicing is a post-transcriptional mechanism that explains how one gene can give rise to multiple protein isoforms with potential different functions, localizations, or stabilities.
A wide variety of human diseases are caused by mutations within cis-acting sequences of pre-mRNAs that produce aberrant alternative splicing and alter function of the mutated allele (Duchenne muscular dystrophy, Cystic fibrosis, Limb girdle muscular dystrophy 1B, among others). In addition, certain splice isoforms are depleted or induced in cancers, diabetes, or muscular dystrophies suggesting specific physiological roles. Defects in the function and/or expression of RNA binding proteins and core spliceosome constituents induce splicing mis-regulation in Myotonic dystrophy, Retinitis pigmentosa, Spinal muscular atrophy, Dilated cardiomyopathy, and Limb girdle muscular dystrophy 1G, among other severe diseases. Tissue specific alternative splicing is critical to maintain tissue identity. Heart and skeletal muscle are two of the tissues where the most tissue specific splicing takes place raising the questions of how developmental stage- and tissue-specific splicing influences protein function and how this regulation takes place. In the lab we are interested on two specific aspects of this big picture question.
1 – How does alternative splicing of trafficking and membrane remodeling genes contribute to the development and maintenance of muscle structure and function?
Striated muscle cells have contractility primordial functions that require a very precise formation of transverse tubules through membrane invagination processes and sarcomere assembly. Receptors and ion channels need to be correctly transported to these locations to exert their key functions in calcium signaling and excitation – contraction coupling. Interestingly, alternative splicing regulates genes involved in trafficking and membrane dynamics specifically in striated muscle development. However, very little is known about the crosstalk between alternative splicing and trafficking/membrane fields. We hypothesize that developmental alternative splicing regulation of genes encoding proteins controlling membrane dynamics and vesicular trafficking allow the expression of specific isoforms required for the maturation and maintenance of the myocyte exquisite architecture.
2 – Which are the roles of epigenetic dynamics on alternative splicing regulation in postnatal striated muscle development?
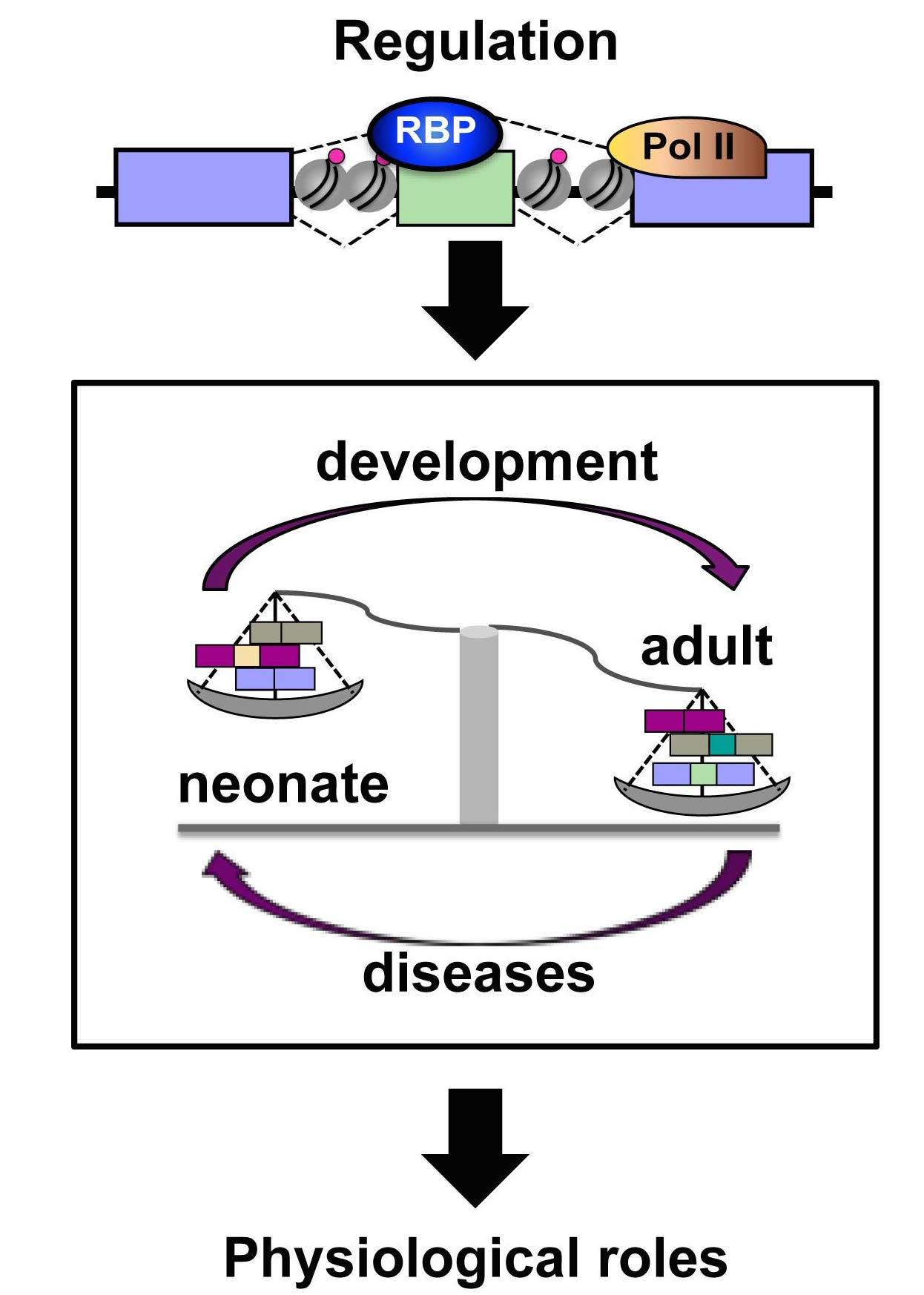
Heart and skeletal muscle development is a very precisely regulated process and chromatin remodelers are essential to suppress the expression of fetal genes in mature myocytes. Damaged adult hearts revert to fetal expression patterns leading to cardiomyopathies, arrhythmias, or congestive failure and chromatin structure changes correlate with this reprogramming. Reversion occurs also in alternative splicing patterns in heart failure and myofiber damage. Although the role of chromatin remodeling on striated muscle gene expression regulation has been extensively studied, its impact on alternative splicing regulation has not been addressed. Our hypothesis is that during muscle development, epigenetic dynamics triggers specific histone marks around the alternative spliced exons altering RNA Polymerase II kinetics, which in turn favors the recognition of weak splice sites and thus inclusion of alternative exons or recruitment of specific splicing factors that promote exon inclusion or skipping.
